

L’invecchiamento, o aging, può anche essere definito come un termine inclusivo per una serie di processi molecolari, biochimici, fisiologici, genetici ed epigenetici che sono stati descritti come i tratti distintivi del processo, decentralizzati e interdipendenti l’uno dall’altro, che portano ad un inevitabile declino funzionale. Meccanismo che va distinto dalla longevità, termine con il quale si vuole più specificamente indicare la capacità di vivere a lungo mantenendo nel tempo una buona qualità della vita, con uno stato di salute fisica, mentale ed emotiva.
Nel corso degli anni, con l’aumento della nostra comprensione del declino biochimico, morfologico e funzionale dei vari processi legati all’invecchiamento, sono in corso sforzi per elaborare strategie e interventi per migliorare gli effetti delle malattie legate all’età. Interventi dietetici come il digiuno intermittente, la restrizione calorica (CR), l’integrazione di mimetici della CR, l’esercizio fisico, ecc., sono stati utilizzati su modelli animali e sull’uomo per rallentare l’invecchiamento, alleviando gli effetti dei processi che rientrano nella categoria dei suoi peculiari segni distintivi. In tale contesto, oltre alle telomerasi, assumono un ruolo cruciale una tipologia speciale di proteine.
Le sirtuine sono una famiglia di istone deacetilasi di classe III dipendenti da nicotina adenina dinucleotide (NAD +), una forma biologicamente attiva della vitamina B3 o niacina, che catalizzano la modificazione post-traduzionale degli istoni e di altre proteine e svolgono un ruolo importante nell’invecchiamento. Le sirtuine dipendono da NAD + per la loro attività e quindi vengono attivate durante condizioni di bassa energia, come il digiuno o l’esercizio fisico. Le sirtuine svolgono un ruolo importante nel metabolismo energetico, nella riparazione del DNA, nell’infiammazione, nella sopravvivenza cellulare, nell’apoptosi, nella senescenza cellulare, nello stress ossidativo e nella produzione mitocondriale e possono essere modulate dalla dieta e dall’esercizio fisico.
Nei mammiferi, sono stati rilevati sette membri della famiglia delle sirtuine, ovvero SIRT1–SIRT7. Sebbene i sette membri abbiano un dominio centrale catalitico altamente conservato identico di 275 amminoacidi, la loro localizzazione all’interno dei compartimenti cellulari varia, con SIRT1, 2, 6 e 7 localizzati nel nucleo, SIRT3, 4 e 5 nei mitocondri e SIRT 1 e 2 nel citoplasma.
Oltre alla deacetilasi, le sirtuine possiedono anche diverse attività enzimatiche, particolarmente cruciali nel contesto della longevità: le proteine bersaglio delle sirtuine includono NF-κB, fattore di trascrizione cruciale pro-infiammatorio e pro-tumorale, l’enzima glicolitico fosfoglicerato mutasi-1 (PGAM-1), la proteina legante l’elemento regolatore dello sterolo (SREBP), il fattore trascrizionale p53, il co-attivatore del recettore attivato dal proliferatore del perossisoma-γ 1α (PGC-1α), la nitrossido sintasi endoteliale (eNOS), mTORC2 e la proteina Forkhead box O3a(FOXO3a), fondamentale per la regolazione cellulare, noto come fattore geroprotettivo legato alla longevità umana.
Tipologie di sirtuine
SIRT1 è la sirtuina dei mammiferi più studiata, specie per il suo ruolo nell’invecchiamento, nell’infiammazione e nella disfunzione metabolica. È localizzata nel nucleo; tuttavia, trasloca nel citoplasma in risposta a determinati stimoli. Svolge un ruolo importante nel riconoscimento e nella risposta al danno del DNA e nel metabolismo attraverso la deacetilazione delle proteine bersaglio che, oltre agli istoni H3, H4 e H1, includono la modifica di diverse proteine non istoniche, inclusi i fattori trascrizionali p53, p65, PGC-1α, NF-κB, SREBP e proteine di riparazione.
SIRT2 è localizzato principalmente nel citoplasma, ma trasloca nel nucleo. Si sposta durante la mitosi e svolge un ruolo importante nel ciclo cellulare.
SIRT3 è una sirtuina mitocondriale che promuove la trascrizione del superossido dismutasi 2 (SOD2) deacetilando e attivando direttamente la SOD2, assumendo un ruolo importante nella protezione antiossidante cellulare. Durante il digiuno prolungato, SIRT3 deacetila l’acil-CoA deidrogenasi a catena lunga (LCAD), una proteina coinvolta nell’ossidazione degli acidi grassi, attivando il processo ossidativo.
SIRT4 è una proteina mitocondriale, priva di attività deacetilasi in vitro. La sovraespressione di SIRT4 è associata ad un aumento dei livelli di ATP e sua la perdita di funzione porta a una diminuzione dei livelli cellulari di ATP in vitro e in vivo.
SIRT5 è un enzima mitocondriale, coinvolto nel metabolismo e nella risposta allo stress, svolgendo un ruolo nel metabolismo energetico mitocondriale. La sovraespressione di Sirt5 aumenta la fosforilazione della proteina chinasi attivata da AMP (AMPK) e attenua la disfunzione mitocondriale, alleviando il danno strutturale mitocondriale e ripristinando le molecole di ATP.
SIRT6 è una sirtuina nucleare. Si è appurato che i topi carenti della proteina SIRT6 mostrano instabilità genomica e invecchiamento precoce. Questo perché SIRT6 mantiene la struttura della cromatina sui telomeri mediante deacetilazione della lisina 9 dell’istone H3, prevenendo il danno al DNA telomerico e la senescenza cellulare. SIRT6 e SIRT1 agiscono sinergicamente in tale contesto.
SIRT7 è un’altra sirtuina nucleare che svolge un ruolo attivo nella biogenesi dei ribosomi, nell’omeostasi metabolica, nella biogenesi mitocondriale, nella riparazione del DNA, nell’infiammazione e nell’apoptosi. È stato riportato che la sovraespressione di SIRT7 nella regione dell’ippocampo nei topi anziani migliora la neurogenesi e le funzioni cognitive.
Fra tutte queste la più studiate sono soprattutto la SIRT1, ma anche sulla SIRT3 e la SIRT6 sono state compiute importanti ricerche che descriviamo in sintesi.
L’effetto della restrizione calorica
Le evidenze più solide indicano che la restrizione calorica (RC) ha un effetto favorente sull’attività e sull’espressione delle sirtuine, sebbene tale effetto sia influenzato dal tipo di restrizione (continua vs intermittente), dal sesso e dal BMI. Uno studio randomizzato (n=112) documenta che sia la RC intermittente che quella continua aumentano la concentrazione di SIRT1 dopo 12 mesi, con l’aumento più marcato osservato negli uomini sottoposti alla RC intermittente.
Analizzando il dettaglio dello studio, nei soggetti con obesità addominale è stato dimostrato che la RC intermittente determina un aumento significativo delle concentrazioni mediane di Sirtuina1 dopo un anno rispetto ai valori basali (da 762 a 884 pg/mL): questo incremento è stato più evidente nei maschi e correlava in modo significativo alle variazioni del BMI nelle donne, mentre negli uomini la correlazione era inversa solo nel gruppo che aveva attuato una RC intermittente.
A sostegno del razionale molecolare, uno studio preclinico molto recente pubblicato sulla rivista Nature suggerisce che la RC porti ad accumulo di acido litocolico (LCA), il quale agisce legandosi a TULP3 (Tubby-like protein 3), un gene che codifica per un fattore di trascrizione cruciale per lo sviluppo embrionale e la funzione neuronale) per attivare direttamente le sirtuine, innescando una cascata che porta all’attivazione dell’AMPK e a un rallentamento dell’invecchiamento.
In sintesi, la restrizione calorica aumenta l’espressione e l’attività delle sirtuine sistemicamente e nei tessuti, con effetti più marcati osservati con la RC intermittente e una risposta influenzata da sesso e BMI. Le sirtuine, in particolare SIRT1, sono attivate anche da mimetici della RC, come il resveratrolo, che potrebbero mediare parte dei benefici anti-aging e metabolici della restrizione calorica, anche se le ricadute cliniche a lungo termine rimangono da chiarire negli studi umani di più ampio respiro. Dal punto di vista clinico, i risultati degli studi suggeriscono che protocolli di restrizione calorica (specialmente intermittente) possono essere considerati come strumento per aumentare l’attivazione sirtuinica, potenzialmente rilevante per la modulazione dell’invecchiamento e della salute metabolica, ma la variabilità dell’effetto rende necessaria una personalizzazione basata su sesso e composizione corporea.
Funzione nelle malattie cardiovascolari
SIRT1 sembra possedere proprietà protettive cardiovascolari, oltre a quelle derivanti esclusivamente dalla forma fisica e metabolica. Ad esempio, SIRT1 e altre sirtuine, possono proteggere dall’ipertrofia delle cellule cardiache e muscolari lisce. SIRT1 protegge anche la muscolatura liscia inibendo l’espressione del recettore AT1 dell’angiotensina. Analizzando ulteriormente l’interazione funzionale tra SIRT1 e SIRT3, la riduzione delle specie reattive dell’ossigeno in modelli sperimentali sembra essere mediata da livelli elevati di SIRT3.
Il primo collegamento tra SIRT1 e le cellule endoteliali è stata la scoperta che SIRT1 deacetila e attiva l’ossido nitrico sintasi endoteliale (eNOS): l’attivazione di eNOS e la repressione di AT1 suggeriscono che l’attività SIRT1 dovrebbe frenare l’ipertensione.
Un’altra funzione di SIRT1 che può essere fondamentale nelle malattie cardiovascolari è la regolazione dell’omeostasi dei grassi e del colesterolo. Oltre a innescare la β-ossidazione degli acidi grassi nella restrizione calorica, SIRT1 esercita anche due effetti opposti sulla sintesi dei grassi e del colesterolo, facilitando il trasporto inverso del colesterolo dai tessuti periferici verso il fegato e, allo stesso tempo aumentando la produzione di lipoproteine HDL (ad alta densità) facilitando la rimozione del colesterolo, esercitando protezione contro l’aterosclerosi.
SIRT 1 e patologie neurodegenerative
Sebbene i metodi per rilevare gli stadi iniziali del morbo di Alzheimer siano diventati sempre più sensibili, gli approcci terapeutici rimangono sfuggenti. Lo stress neuronale (valutato, ad esempio, nei neuroni in coltura o nei topi transgenici) è stato mitigato dalla sovraespressione di SIRT1, suggerendo la questione se questa sirtuina potrebbe frenare la malattia di Alzheimer.
Infatti, è stato dimostrato che la sovraespressione di SIRT1 nel cervello riduce il carico del peptide β-amiloide, l’agente tossico generato dalla scissione proteolitica della proteina precursore dell’amiloide nei topi che sovraesprimono due geni umani predisponenti alla malattia di Alzheimer precoce. SIRT1 è capace di attivare il gene che codifica per l’α-secretasi, enzima proteolitico che scinde la proteina precursore dell’amiloide, impedendo la formazione di placche tossiche di peptide β-amiloide e quindi proteggendo dalla malattia. SIRT1 ha attivato il gene dell’α-secretasi deacetilando il suo attivatore trascrizionale, il recettore β dell’acido retinoico. A conferma di ciò, nei topi che sovraesprimono il peptide β-amiloide, il livello di placche è decisamente ridotto in quelli che sovraesprimono anche SIRT1.
Composti naturali attivatori
Il resveratrolo è uno dei primi composti scoperti come sostanza mimetica della restrizione calorica nell’attivare le sirtuine. Si trova in piante come uva, mirtilli, mele, prugne, arachidi e prodotti a base di uva come il vino rosso. Interagisce con un’ampia varietà di proteine che fungono da messaggeri cellulari, tra cui Nrf2 e le sirtuine. Il resveratrolo modula anche l’attività deacetilasica di SIRT3 e SIRT5, oltre a regolare positivamente l’espressione dei geni che codificano gli antiossidanti e dei fattori di neuroprotezione.
Si verifica una stretta interazione tra i sensori energetici, AMPK e SIRT1; infatti, il resveratrolo li regola in modo dose-dipendente e reciproco.
La curcumina agisce anche come attivatore di SIRT1, come pure anche di altre sirtuine. La down-regulation di SIRT1 dopo l’infarto del miocardio è stata attenuata dal pretrattamento con curcumina, suggerendo che l’attivazione di SIRT1 potrebbe essere attivata dalla curcumina. Il pretrattamento con curcumina, sempre mediante l’attivazione di SIRT1, protegge anche da una serie di disturbi neurologici come l’ictus, oltre a bloccare la neurotossicità della β-amiloide nei neuroni corticali del ratto. Un aumento del livello/attività di AMPK e SIRT1 è stato osservato nei muscoli di topi e ratti, dopo l’integrazione di curcumina, migliorando l’effetto dell’esercizio.
Il pterostilbene è un analogo (un dimetil etere) del resveratrolo, sicuro per il consumo umano con un’elevata biodisponibilità di cui sono ricchi i mirtilli neri. In studi recenti, è stato scoperto che il pterostilbene attiva le sirtuine, l’AMPK e l’Nrf2. In un modello di topi obesi, questo composto migliora la termogenesi e la biogenesi mitocondriale attivando il percorso Sirt1/PGC-1α/Sirt3.
Niacinamide (Vitamina B3), come vitamina è essenziale per oltre 500 reazioni enzimatiche nel nostro organismo e fondamentale per quasi tutti i processi biologici. Numerosi studi hanno evidenziato il suo ruolo chiave extra vitaminiconella longevità cellulare: la sua supplementazione ha mostrato la capacità di inibire la calcificazione vascolare attraverso il mantenimento dei livelli sia di SIRT1 che di SIRT6, dimostrandosi un potente attivatore della longevità cellulare riducendo anche l’autofagia.
Come inizialmente ricordato, l’attività delle sirtuine è strettamente dipendente dal NAD +, per il quale la niacinamide ne contribuisce all’aumento dei livelli, essendo una sua forma derivata. Studi recenti indicano che l’assunzione di niacinamide, anche in combinazione con i suoi derivati, aumenta in modo dosedipendente la concentrazione di NAD +, potenziando i meccanismi cellulari associati alla longevità e alla protezione contro l’invecchiamento.
Esistono altri composti naturali noti per attivare le sirtuine. Questi includono flavonoidi come fisetina, quercetina, apigenina, epigallocatechina gallato del tè verde (EGCG) e diverse altre.
Nei topi, è stato scoperto che l’honokiolo estratto dalla magnolia inverte l’ipertrofia cardiaca attivando SIRT3. La quercetina e i suoi derivati, ad alte concentrazioni, attivano SIRT6 legandosi al suo canale di legame acilico selettivo; anche i polifenoli come la cianidina e la luteolina attivano questa sirtuina.
Infine, la fisetina e la luteolina sopprimono lo stress ossidativo attivando le sirtuine come SIRT1, SIRT3 e SIRT6 e riducendo al tempo stesso l’espressione di FOXO3a.
Le istone deacetilasi (HDAC)
Le istone deacetilasi (HDAC) sono enzimi che regolano l’espressione genica rimuovendo i gruppi acetilici dalle lisine sulle code degli istoni, aumentando la carica positiva delle code istoniche, condensando e compattando la cromatina (da eucromatina a eterocromatina).
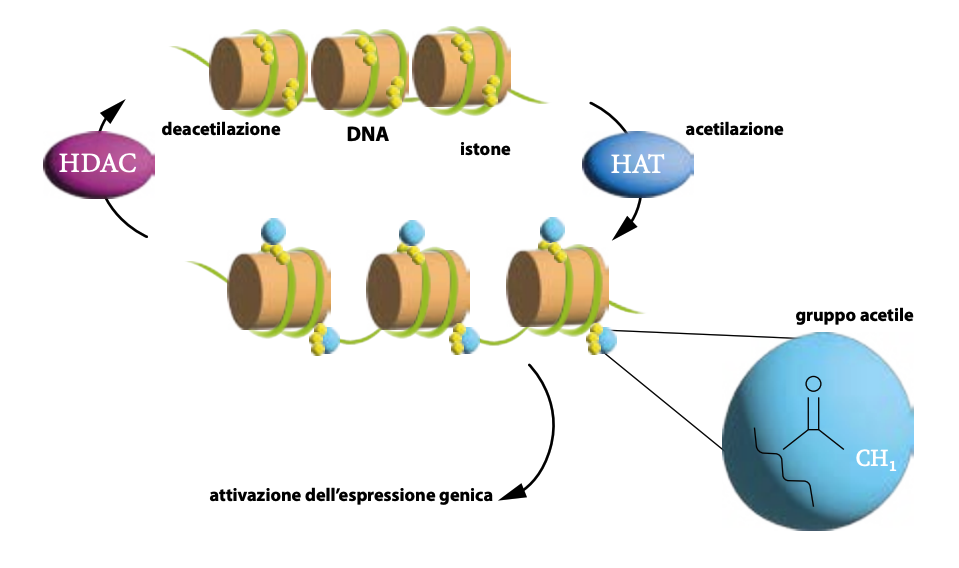
Questa azione aumenta l’affinità tra istoni e DNA, rendendolo meno accessibile ai fattori di trascrizione e provocando il silenziamento genico. Regolano proliferazione, differenziazione e apoptosi cellulare. Sono cruciali in processi biologici come il ciclo cellulare, l’apoptosi e lo sviluppo di patologie come cancro e malattie neurodegenerative.
L’AMPK
L’AMPK è un enzima regolatore chiave che monitora lo stato energetico cellulare e si attiva in risposta a bassi livelli di energia, svolgendo un ruolo significativo nel mantenimento dell’equilibrio energetico all’interno delle cellule.
L’uso degli attivatori di AMPK consente di studiare i meccanismi con cui le cellule percepiscono e rispondono ai cambiamenti nella disponibilità di energia, nonché di esplorare le vie coinvolte nei processi metabolici come l’assorbimento del glucosio, l’ossidazione degli acidi grassi e la biogenesi mitocondriale.
Questi attivatori sono fondamentali per analizzare il ruolo dell’AMPK in vari contesti fisiologici, tra cui le risposte allo stress, la fisiologia dell’esercizio fisico e il rilevamento dei nutrienti.
Tratto dal numero di giugno 2026 di Medicina Integrata
Bibliografia
- López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: An expanding universe. Cell. 2023;186(2):243-278.
- Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403(6771):795-800.
- Bonkowski MS, Sinclair DA. Slowing ageing by design: the rise of NAD+ and sirtuin-activating compounds. Nat Rev Mol Cell Biol. 2016; 17:679-90.
- Zhao L, Cao J, Hu K, He X, Yun D, Tong T, et al. Sirtuins and their Biological Relevance in Aging and Age-Related Diseases. Aging Dis. 2020; 11:927-45.
- Haigis, M. C., and Sinclair, D. A. Mammalian sirtuins: biological insights and disease relevance. Annu. Rev. Pathol. 2010; 5, 253–295.
- Chen D, Steele AD, Lindquist S, Guarente L. Increase in activity during calorie restriction requires Sirt1. Science. 2005;310(5754):1641.
- Donmez G, Outeiro TF. SIRT1 and SIRT2: emerging targets in neurodegeneration. EMBO Mol Med. 2013; 5:344-52.
- Michan S, Sinclair D. Sirtuins in mammals: insights into their biological function. Biochem J. 2007;404(1):1-13.
- Wood JG, Rogina B, Lavu S, Howitz K, Helfand SL, Tatar M, et al. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature. 2004; 430:686-9.
- Cantò C, Gerhart-Hines Z, Feige JN, Lagouge M, Noriega L, Milne JC, et al. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature. 2009; 458:1056-60.
- Borra MT, Smith BC, Denu JM. Mechanism of human SIRT1 activation by resveratrol. J Biol Chem. 2005;280(17):17187-17195.
- Zendedel E, Butler AE, Atkin SL, Sahebkar A. Impact of curcumin on sirtuins: A review. J Cell Biochem. 2018; 119:10291-300.
- Li YR, Li S, Lin CC. Effect of resveratrol and pterostilbene on aging and longevity. Biofactors. 2018;44(1):69-82.
- Mills KF, Yoshida S, Stein LR, Grozio A, Kubota S, Sasaki Y, et al. Long-Term Administration of Nicotinamide Mononucleotide Mitigates Age-Associated Physiological Decline in Mice. Cell Metab. 2016; 24:795-806.







{kind=link}